
| |
| Volume 15, Number 4 | September 18, 2025 |
In the first installment of this series, I recommended an intake of 500 mg NR (nicotinamide riboside) per day for users of Dr. Aardsma's Anti-Aging Vitamins over the age of 40 years.[1] While NR has unquestionably been helpful in combating post-aging disease, a better alternative is now on the horizon. I am now recommending cessation of NR supplementation in anticipation of this better alternative. The present article explains the rationale behind this recommendation.
The hypothesis prompting my investigation into NR supplementation was formulated in the first part of this series as follows:[2]
NAD Destruction Hypothesis: In the absence of dietary MePiA, ROS-damaged mitochondria deliberately destroy NAD to slow further ROS production via oxidative phosphorylation to limit the rate of further ROS damage.
This now requires amending. The words "destroy NAD" need to be replaced with "limit NAD concentration." The hypothesis then reads:
NAD Destruction Hypothesis: In the absence of dietary MePiA, ROS-damaged mitochondria deliberately limit NAD concentration to slow further ROS production via oxidative phosphorylation to limit the rate of further ROS damage.
The need for this amended reading results from the fact that the NADase enzyme, CD38, previously discussed in this series, is now seen as not only functioning to destroy NAD+ but also to destroy NAD+ precursors.
However, we and others demonstrated that CD38 degrades not only NAD, but also circulating [in the blood] NAD precursors such as NMN and NR, before they can be incorporated inside cells for NAD biosynthetic pathways.[3]This, as it turns out, implies that CD38 destruction of NAD+ in blood plasma is only incidental. This incidental destruction has very little impact on the level of NAD+ within cells. This incidental destruction is not how ROS-damaged mitochondria limit NAD+ concentrations inside cells. Rather, intracellular NAD+ concentration is limited by preventing NAD+ precursors from ever reaching the cells.
NAD+ inside cells may be thought of as being limited via the strategy of siege warfare. Blood plasma is the highway on which nutrients are transported to cells. The highway is populated with CD38 troops, called out to protect the country (the body) from destruction due to ROS damage. To accomplish their mission, the troops destroy NAD+ precursors found traveling along the highway. They never manage to destroy 100% of their targets. But, as a result of their destructive labors, cells are slowly starved of NAD+.
Cells are conservative of NAD+. They recycle used NAD+ via so-called salvage pathways internal to the cell. So they do not immediately run out of NAD+. But, by limiting the production of new NAD+ within cells, it is inevitable that cells will slowly become deficient in NAD+.
Thus the original hypothesis required correction. It was built on the mistaken idea that, by increasing the abundance of CD38, more NAD+ gets destroyed, reducing the availability of NAD+ for oxidative phosphorylation (in the mitochondria) with its accompanying harmful ROS production. The amended hypothesis corrects this. It is built on the idea that, by increasing the abundance of CD38, less NAD+ gets created in cells, reducing the availability of NAD+ for oxidative phosphorylation (in the mitochondria) with its accompanying harmful ROS production.
This modifies our NAD+ flow and distribution model once again.
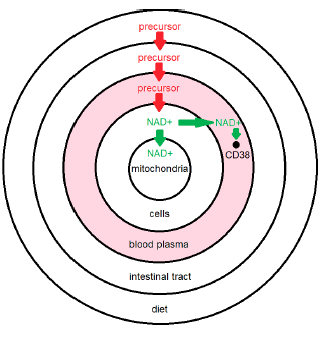
The previous model (Figure 1), when corrected, yields the model shown in Figure 2. This figure immediately clarifies several things.
 |
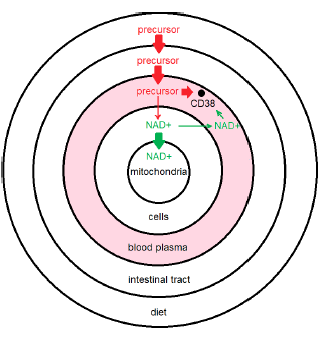 |
First, it shows that loss of NAD+ from blood plasma is incidental. This loss plays only a minor role in the maintenance of necessary NAD+ concentrations in cells. The main thing going on in blood plasma having real consequence for NAD+ concentration in cells is removal of dietary precursors of NAD+, such as NR, limiting the availability of these precursors to cells for replenishment of intracellular NAD+.
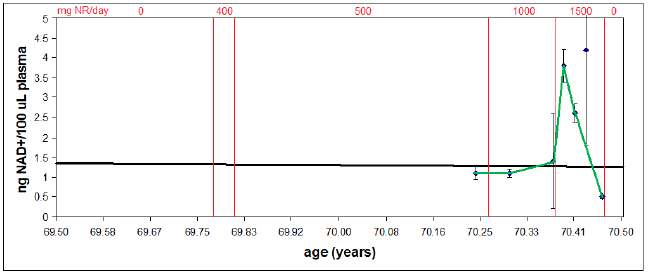
Second, it clarifies why reduction of my blood plasma NAD+ levels (shared last issue and reproduced here as Figure 3) to near zero, due to overshooting the mark with NR supplementation, did not result in any accompanying health issues—why I didn't suddenly feel 90 years old. Low NAD+ levels in blood plasma are incidental to CD38 overexpression. They are not causative of disease. They are symptomatic of disease. I will call this disease "CD38 overexpression disease." It is discussed further below.
 |
Third, it shows that there is, nevertheless, an incidental loss of NAD+ from blood plasma. This is fortunate as it facilitates measurement of the extent of CD38 overexpression. When CD38 is being properly expressed, youthful levels of blood plasma NAD+ will be the result. Thus the level of NAD+ in blood plasma allows us to see whether different CD38 modulation strategies are succeeding.
Fourth, it clarifies the interpretation of my blood plasma NAD+ level measurements made while supplementing NR.
Initially, despite many months of supplementation with NR, the green curve in Figure 3 shows that my NAD+ level remained normal for my age. This is now explained by the new model as due to the destruction of most NR in the blood plasma by CD38.
The CD38 sink was finally overwhelmed when I increased my daily intake from 1,000 to 1,500 mg NR. This resulted in a rapid increase in my plasma NAD+, restoring it to youthful levels. This indicates that the cells domain was similarly being restored to youthful levels since the model shows that there is no way for NAD+ to increase in plasma apart from resupply by the cells. This conclusion is supported by a simultaneous return to a more youthful physiology, most noticeable in regard to sexual potency but also noticeable with reduction in general aches and pains.
Unfortunately, this rapid increase was followed by nearly as rapid decline. This decline is explained as follows.
Resupply of NAD+ to the cells during the rapid increase revved up oxidative phosphorylation in the mitochondria, which was the desired effect. But 1,500 mg NR turned out to be excessive, producing excessive oxidative phosphorylation. The result was excessive ROS generation—that is, ROS generation exceeded the level which could be handled by available MePiA molecules in the mitochondria. This excessive ROS caused biomolecular damage to the mitochondria, triggering production of more CD38, as it always does, enlarging the CD38 sink. This enlarged sink caused rapid decline in my plasma NAD+, to levels typical of individuals in their 90s.
This reduction in plasma NAD+ levels does not appear to pertain in the cells domain since a more youthful physiology still pertains at present. Thus, the plasma decline is indicative only of an enlarged CD38 sink and not of NAD+ deficiency in the cells domain.
The big negative, from a theoretical perspective, is that it will be more difficult to modulate the CD38 sink going forward, since it has been made larger than it was prior to NR supplementation.
Finally, the new model shows immediately a weakness with the strategy of boosting NAD+ levels by the addition of dietary precursor. It shows that most NAD+ precursor simply goes down the drain. Dietary precursor is largely eliminated by the CD38 sink, with little surviving to do any good in the cells where it is needed. In the least case, this is bad economically. Most of the dietary supplement NR we have been paying for has not benefited us.
But there is another potential downside with more serious potential consequences. CD38 functions not only as an enzyme but also as a signaling molecule, regulating a number of cellular processes. Thus the enlarged (overexpressed) CD38 sink clearly has potential to upset these other processes, creating further health problems.
Clearly, NAD+ deficiency is not the fundamental problem. It is a derivative problem which was created by the overexpression of CD38 all the years during which our diets contained no vitamin MePiA. The fundamental problem, remaining even after MePiA has been restored to the diet, is continued presence of too much CD38.
It is this fundamental problem—CD38 overexpression disease—which needs to be targeted for correction for those of us now supplementing our diets with Dr. Aardsma's Anti-Aging Vitamins. Supplementation with NR, though helpful for boosting NAD+ in besieged cells, fails to address this fundamental problem. When CD38 overexpression disease has been properly dealt with, the siege will be lifted, yielding the expectation that NAD+ deficiency disease will then disappear without any need for NR supplementation because other natural dietary precursors (such as vitamin B3) will then be able to get through to the cells.
Though dietary supplementation with NR made progress against post-aging, aging-induced disease, it failed to target the fundamental problem of aging-induced CD38 overexpression disease. I have, for this reason, stopped dietary supplementation of NR for Helen and myself, and I recommend that users of Dr. Aardsma's Anti-Aging Vitamins do the same.
I have now begun to research what might be done to address CD38 overexpression disease using flavonoid inhibitors of CD38. Flavonoids are natural compounds derived from plants. I will, of course, keep you posted in these pages as I learn more.
Having made aging my highest research priority for a quarter of a century now, and having solved aging disease itself,[4] it currently appears that we may, at long last, be nearing a reasonably complete solution to the ravages of the syndrome commonly called "old age" today. ◇
The Biblical Chronologist is written and edited by Gerald E. Aardsma, a Ph.D. scientist (nuclear physics) with special background in radioisotopic dating methods such as radiocarbon. The Biblical Chronologist has a fourfold purpose:
The Biblical Chronologist (ISSN 1081-762X) is published by: Aardsma Research & Publishing Copyright © 2025 by Aardsma Research & Publishing. |
^ Gerald E. Aardsma, "Dietary Supplementation with NR for the Post-aging Diet," The Biblical Chronologist 14.17 (December 12, 2024): 7. www.BiblicalChronologist.org.
^ Gerald E. Aardsma, "Dietary Supplementation with NR for the Post-aging Diet," The Biblical Chronologist 14.17 (December 12, 2024): 5. www.BiblicalChronologist.org.
^ E.N. Chini et al., "The Pharmacology of CD38/ NADase: An Emerging Target in Cancer and Diseases of Aging" Trends in Pharmacological Science 39.4 (2018): 424–436. pmc.ncbi.nlm.nih.gov/articles/PMC5885288/.
^ Gerald E. Aardsma, Aging: Cause and Cure, 3rd ed. (Loda, IL: Aardsma Research and Publishing, 2023). www.BiblicalChronologist.org.