
| |
| Volume 14, Number 17 | December 12, 2024 |
Helen and I have now added nicotinamide riboside (NR) supplementation to our daily diets. This is in addition to daily supplementation with Dr. Aardsma's Anti-Aging Vitamins.
Briefly, supplementation with Dr. Aardsma's Anti-Aging Vitamins cures aging, but it does not cure aging-induced mitochondrial microheteroplasmy disease. This disease causes ongoing slow health decline via energy starvation of cells even after aging has been cured. Theory indicates that supplementation with NR should mitigate energy starvation due to microheteroplasmy and thereby facilitate return to a more youthful state of health.
The purpose of the present article, at this very early stage of my research into supplementation with NR, is to show in detail why and how we are adding NR to our diets to make it as easy as possible for others who are experiencing the negative impacts of microheteroplasmy disease to follow our lead.
The third edition of Aging: Cause and Cure detailed the state of knowledge, up to 2021, stemming from Bible/science research into aging.[1] It explained modern human aging as follows.
Theory of Modern Human Aging: Modern human aging is a congenital nutritional deficiency disease syndrome (called MHA) of two newly-discovered, closely-related vitamins: methylphosphinic acid (MePiA) and methylphosphonic acid (MePA).
Vitamin MePiA functions as an antioxidant within the mitochondria, protecting them from free radical damage due to reactive oxygen species (ROS). The fundamental cause of death resulting from vitamin MePiA deficiency is cellular energy starvation due to decreased energy output from ROS-damaged mitochondria.
Vitamin MePA functions as its own unique vitamin, separate from vitamin MePiA. It is involved in multiple biochemical pathways, similar to members of the traditional vitamins, and like them its dietary deficiency gives rise to its own unique, potentially fatal disease.
Since that time, while a few (4) of these BC research newsletters have dealt with further progress in aging research, the large majority (25) have been concerned with Bible/science research into other matters. This reflects deliberate research strategy.
Aging research is given highest priority with me because of its potential to alleviate human suffering and bereavement. But further theoretical advance in regard to aging was stalled following publication of Aging: Cause and Cure, third edition, while I was waiting on experimental results.
It was clear that the damage done to the body by MePA deficiency disease healed relatively quickly once dietary supplementation with MePA had begun. MePA deficiency disease seemed then, as it still does today, to have been conquered.
But not so the aftermath of MePiA deficiency disease. The big question on which I needed experimental input was whether mitochondrial microheteroplasmy resulting from MePiA deficiency disease might also heal itself in a reasonably short time, once dietary supplementation with MePiA had begun. If it did, further research into how to combat microheteroplasmy would be a waste of effort, time, and money.
So I waited, turning back to the unfinished Bible/science problem of the route of the Exodus, and turning also to the previously untackled Bible/science problem of the cause of Earth's rapidly changing climate and modern global warming.
Both of these problems have now been solved,[2] opening opportunity to turn back to the problem of microheteroplasmy once again.
I was never sanguine about the likely rate of healing of microheteroplasmy. The following lengthy quote from the third edition of Aging: Cause and Cure shows this while also providing some helpful review.[3]
Anecdotal evidence from users of Dr. Aardsma's Anti-Aging Vitamins, coupled with close-up observations of Helen and myself (also anecdotal evidence), plus early results from several mouse experiments currently underway have now confirmed that cellular energy starvation due to microheteroplasmy does not resolve quickly on its own.The healing of the mess made by MePiA deficiency disease, with its consequent energy starvation of cells, is a more complex matter. Once adequate daily intake of MePiA has begun, further ROS damage to mitochondria is greatly attenuated. The [computer] program approximates this, somewhat optimistically again, by reducing further ROS damage immediately to zero. This treats MePiA deficiency disease as immediately cured, which seems a reasonable approximation because there is no longer any deficiency of MePiA and further wildfire ROS damage has been halted. MHA has been cured by beginning to take the anti-aging vitamins, but full health has not yet been restored. Stopping further wildfire ROS damage has not erased the damage already done to the mitochondria.
Present theory says that it is mutation of the mtDNA due to ROS damage which is responsible for declining mitochondrial energy production. Individual copies of the mtDNA have been mutated in divergent ways, seeding heteroplasmy throughout the organism. This sort of heteroplasmy is called "microheteroplasmy." Microheteroplasmy is the damage already done.
Microheteroplasmy is not aging, and it is not an aging disease. It is, like cancer, an aging-related disease. It is its own disease, induced by the aging disease called MePiA deficiency disease. Curing aging does not automatically cure all aging-related diseases. Curing MHA does not automatically cure microheteroplasmy.
The damage already done—microheteroplasmy—represents not simply harm to some biomolecule which might easily be repaired, but rather a loss of information from the mitochondrial blueprints regarding what the biomolecules needed by the organism are meant to look like. Restoration of this lost information is a difficult problem for the organism to solve at the biomolecular level. The mitochondrion does not "know" what undamaged mtDNA looks like. The mitochondrion possesses no indelible mtDNA copy which can be referenced in order to distinguish between pristine mtDNA and mutated mtDNA. The mitochondrion is merely a mindless machine, pumping out whatever biomolecules its multiple mtDNA copies—pristine or otherwise—specify. It appears that the mitochondrion has no way of fixing itself—no way of restoring its lost information.
It thus appears that damaged mitochondria stay damaged. But this is not the end of the story. There still seems to be a possibility of at least some degree of healing.
Theoretically, it seems possible for the organism's cells to discriminate against mitochondria that are less efficient because they have been damaged, replacing them with new copies of mitochondria that are more efficient because they are less damaged. This does not immediately restore pristine mitochondria, but it does begin to clean up the microheteroplasmy mess.
Theoretically, this can eventually restore pristine mitochondria. In the process of generating new mitochondria, by fission of a less damaged mitochondrion, mtDNA copies from the parent mitochondrion would be expected to be randomly apportioned between the two offspring mitochondria. This would result in offspring mitochondria with differing energy-production efficiencies. If, for example, one offspring mitochondrion happened to get all of the best-preserved mtDNA copies from the parent, while the other got all the most-damaged copies, this would allow further refining of the mtDNA via further discrimination against the less-efficient offspring mitochondrion.
Even if the body is able to do this, it is expected to be a very slow process, lasting decades.
The time has clearly come for a deliberate assault on this problem.
There are two obvious, categorically different research strategies for achieving mitigation of microheteroplasmy disease.
The first is to edit the mtDNA to restore its original blueprint. This is a very tall order.
The body contains on the order of 10 quadrillion mitochondria, and on average, each mitochondrion possesses on the order of 1,000 copies of mtDNA. Microheteroplasmy involves not just a single mutation common to all copies, but rather multiple, varied, random mutations to each and every one of these copies—a monstrous mess.
Nonetheless, mitochondrial gene editing is beginning to be tackled by medical science, and it seems probable to me that it will ultimately become possible to clean up this microheteroplasmy mess therapeutically. However, the technology which is needed to tackle this problem is still being developed, and we very much need something which can be done now.
This brings us to the second strategy.
The second strategy does not seek to restore the damaged mitochondria, but rather to restore to the body whatever molecules may be missing because of the damaged, dysfunctional mitochondria.
This approach is familiar to modern medicine. For example, for those of us with worn-out, dysfunctional thyroid glands, modern medicine typically uses a daily levothyroxine supplement—a pill—to provide the body with what the thyroid gland is no longer able to produce in adequate amount.
The major role of the mitochondria is to supply cells with energy. The energy molecule for cells is adenosine triphosphate (ATP). Thus it would seem, at first glance, that supplementing ATP might be the way to go. But this does not appear to be a good solution to microheteroplasmy for two reasons.
First, ATP is not adsorbed by the cells when taken orally. This has been demonstrated by deliberate measurements of ATP in the blood of eight healthy volunteers following oral administration of a hefty intake (5000 mg) of synthetic ATP.[4]
One could imagine intravenous administration of ATP, but this would not be very practical in regard to time and expense. Second, when due consideration is given to what appears to be going on in the body leading to energy starvation of the cells (discussed in the next section), any attempt at lifelong ATP supplementation seems likely to end badly. The reason for this is that the biochemical reactions which produce ATP spin off other biological molecules which are vital to cells. One would really rather figure out how to revitalize these ATP-producing reactions so that production of these spinoff molecules is at the same time revitalized.
Astonishingly, this more subtle version of Strategy 2 appears to be possible. Revitalizing ATP-producing reactions seems to be possible by boosting physiological levels of nicotinamide adenine dinucleotide (NAD), which can be done by dietary intake of NAD precursors such as NR.
Before rushing off in this direction, however, it is vital to understand what is going on with NAD in the body.
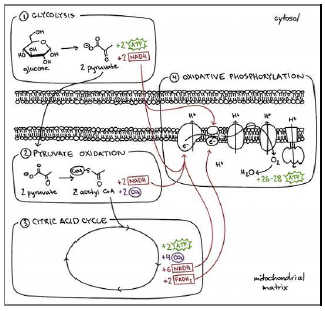
In the body, NAD exists in two forms, designated NADH and NAD+. NAD is intricately associated with production of ATP (Figure 1). As the human body ages, NAD declines (Figure 2). It is most important to understand that NAD does not decline incidentally with age. Rather, it is increasingly targeted for destruction.[5] Why should this be the case?
 |
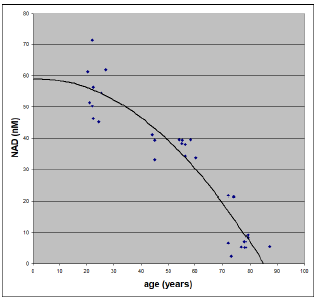 |
I suggest that NAD is targeted for destruction as a defense mechanism against ROS damage in the absence of dietary MePiA. The Figure 2 data pertain, of course, to individuals whose diets are completely deficient in MePiA. For individuals having an adequate daily intake of MePiA all their lives, such as individuals living before Noah's Flood, this graph from birth to 100 years of age would show a constant rather than a falling NAD concentration.
Figure 1 shows how cells produce the energy they need. The MHA theory of human aging focuses attention on the oxidative phosphorylation compartment. It does so for three reasons:
This compartment is pretty much what mtDNA is all about.
So, altogether mtDNA encodes for two rRNAs, 22 tRNAs, and 13 protein subunits, all of which are involved in the oxidative phosphorylation process.[6]This means that ROS-damaged, mutated mtDNA will hinder energy production in this compartment and only in this compartment.
This is the high-energy-output compartment. Loss of function in this compartment and only in this compartment explains the saturating nature of energy starvation of cells observed via modeling of the Social Security Administration's actuarial life table data.[7]
This compartment, with its high-energy, oxygen reactions, is the heart of where unwanted, toxic ROS are generated.[8] Thus, this compartment is where MePiA is meant to be, and where, when present, MePiA acts to keep ROS from damaging mtDNA and other biomolecules throughout the mitochondria.
In the absence of dietary MePiA, ROS damage rages out of control within the mitochondria. To limit this damage—in the absence of MePiA—it is necessary to limit oxidative phosphorylation. This can be achieved by limiting availability of NAD, which limits its NADH form, which is, as the figure shows, the main "fuel" feeding the oxidative phosphorylation "fire." To this end, NAD is destroyed by increasing levels of "CD38," a "NADase" molecule, which breaks NAD down into smaller molecules.
In the absence of dietary MePiA, the body limits NAD because the body is trying to protect itself from out-of-control ROS. To let ROS generation carry on unchecked is to wind up soon dead due to destruction of the energy producing oxidative phosphorylation apparatus together with the mtDNA blueprints for how to make new copies of that apparatus, with inevitable severe energy starvation the result. To shut down oxidative phosphorylation completely, so as to stop out-of-control ROS damage, is to wind up with this same outcome—severe energy starvation and death. There is clearly an optimum for longevity somewhere between these two extremes. MePiA-deprived mitochondria auto-regulate oxidative phosphorylation via NAD destruction to achieve this optimum.
Because this deliberate-destruction concept appears crucial to proper use of NAD supplementation, I will highlight it by naming it the NAD Destruction Hypothesis:
NAD Destruction Hypothesis: In the absence of dietary MePiA, ROS-damaged mitochondria deliberately destroy NAD to slow further ROS production via oxidative phosphorylation to limit the rate of further ROS damage.
The downside of limiting oxidative phosphorylation is that less ATP gets produced. This means less energy available for cells than needed. Running the body with low cellular energy levels results in cells having insufficient energy to do all of the many tasks they were meant to do. Some important tasks will be left undone because the cells lack the energy needed to carry them out. Health and longevity will necessarily suffer. The organism will slowly decline. Thus, restoring youthful energy levels to cells is vitally important.
The interval of time which has elapsed since publication of the third edition of Aging: Cause and Cure has been sufficient to reveal that cells do not naturally recover quickly, if at all, from reduced cellular energy levels due to microheteroplasmy disease. Intervention appears necessary. Boosting NAD via supplementation with NR appears to be a functional intervention.
But the NAD Destruction Hypothesis implies that boosting NAD via supplementation with NAD precursors in the absence of MePiA is not a good idea. This will rev up oxidative phosphorylation, increasing cellular energy levels, but it will at the same time rev up production of toxic, destructive ROS (which is why the body is limiting NAD in the first place). The immediate result may be improved energy levels, but the long-term outcome can only be shortened life expectancy due to accelerated ROS damage.
To boost NAD safely, MePiA absolutely must be present in the diet together with a NAD precursor. In the presence of MePiA, ROS are no longer out of control. And in that case, boosting NAD back up to youthful levels to improve energy availability and overall health makes a great deal of sense because of the numerous biochemical processes in which NAD is involved, not just the processes of energy production.
Said simply, for those of us who have suffered advanced aging disease, supplementing our diets with a NAD booster in addition to Dr. Aardsma's Anti-Aging Vitamins appears to be a safe, effective, and most welcome way to alleviate microheteroplasmy disease.
There are a number of commercially available NAD precursor dietary supplements. The body processes these precursors to yield NAD, boosting NAD levels in the blood, which makes increased NAD levels available to cells.
The simplest precursor is the well-known vitamin B3, also known as niacin. It does not work out well as a supplement for boosting NAD because it has a relatively low upper limit for daily intake—just 35 milligrams per day. This upper limit has been set as a result of long medical experience with negative side effects experienced by individuals taking too much niacin.
NR does the same thing as vitamin B3 but without the negative side effects.
After overnight fasting, a healthy 52-year-old male self-administered 1,000 mg of NR Cl [i.e., NR] orally at 8 am on 7 consecutive days. Blood and urine were collected for quantitative NAD+ metabolomic analysis. The participant took 0.25 g of NA [i.e., 250 mg of nicotinic acid, also known as vitamin B3 or niacin] to assess sensitivity to flushing and self-reported painful flushing that lasted 1 h. No flushing was experienced on NR.[9]
Typical daily intakes for NR supplementation capsules are in the range of 100 to 500 mg. Many commercial NR supplements include in the capsule other nutraceuticals such as resveratrol and trimethylglycine. My choice is not to use NR which has been combined with any extra nutraceutical ingredients. I make this choice for both theoretical and practical reasons.
From the theoretical side, so far, MHA theory finds no place for any of them. I am not trying to do random experiments with my highly complex body. I am trying to be rational—to understand my body's complexity and then act intelligently to obtain the best outcome for health and longevity. We have seen above that theory indicates that taking even just NR by itself, in the absence of Dr. Aardsma's Anti-Aging Vitamins, will be harmful long term, though many people are doing so ignorantly and innocently at present. It is easy to go wrong. Prudence says to tune out the hype surrounding the fads and stick to a rational strategy.
From the practical side, adding unnecessary extras complicates the problem of side effects. Each extra ingredient increases the possibility of unwanted side effects, and the more extra ingredients there are, the more difficult it is to figure out what is doing what. It makes best sense to keep things as simple as possible right from the start.
This reasoning extends also to the form of NR itself. Commercially available NR seems to be widely available in two forms: 1) NR chloride (NR Cl), and 2) NR hydrogen malate. Look on the nutrition label. NR chloride is the natural form. If the nutrition label says "Nicotinamide Riboside" or "Nicotinamide Riboside Chloride," then it is NR chloride. If it says "Nicotinamide Riboside Hydrogen Malate," then it is NR hydrogen malate. The hydrogen malate form is easier to manufacture and purify, which makes it less expensive to produce, but hydrogen malate comes from malic acid, which, though naturally present in many foods, has its own list of potential side effects. So my choice is to go with the NR chloride form.
After a great deal of searching, Helen was able to locate an NR-only product for us which contains 500 mg of nothing but NR per capsule.[10]
At present, Helen and I take one NR capsule every morning at breakfast together with our daily intake of Dr. Aardsma's Anti-Aging Vitamins.
I can share at this point that NR appears to have been quickly effective with me. After taking my first capsule at breakfast, I went to work in my home office. An hour or two later, deep in thought, I rose from my chair—and my thinking was immediately interrupted by the thought, "That was easy!" I meant that it was easier than usual getting up out of my chair—as though I had suddenly gotten lighter. I hadn't gotten lighter, of course. My legs had gotten stronger. This has subsequently been verified by ease of climbing stairs and reduced fatigue following our daily two-mile walk. I have also noticed reduced brain fatigue, which is especially welcome.
Helen has always had high physical energy, so NR has made little difference in regard to physical fatigue with her. But she, too, is finding reduced brain fatigue, especially noticeable from mid-afternoon until bedtime. And she is noticing improved sleep once again, something she noticed a dramatic improvement in when she first began supplementing MePA some years ago.
For users of Dr. Aardsma's Anti-Aging Vitamins, theory plus limited experience imply that dietary supplementation with pure NR affords a means of compensating for ROS-damaged, dysfunctional mitochondria, improving health and longevity. At this early stage of investigation, I recommend including NR at 500 mg per day in the diet of anyone over the age of forty who uses Dr. Aardsma's Anti-Aging Vitamins.
Let me emphasize, before closing, that I do not recommend supplementing with NR at any level in the absence from the diet of Dr. Aardsma's Anti-Aging Vitamins as theory indicates that this will result in long-term harm to health and longevity.
I expect to be working out a lab procedure for monitoring NAD levels so daily intake of NR can be adjusted: 1) to optimize NAD concentration in blood plasma, and 2) to minimize NR supplementation expense. This is expected to refine this simple 500-mg-per-day initial recommendation. I will, of course, share what I eventually learn. ◇
The Biblical Chronologist is written and edited by Gerald E. Aardsma, a Ph.D. scientist (nuclear physics) with special background in radioisotopic dating methods such as radiocarbon. The Biblical Chronologist has a fourfold purpose:
The Biblical Chronologist (ISSN 1081-762X) is published by: Aardsma Research & Publishing Copyright © 2024 by Aardsma Research & Publishing. } |
^ Gerald E. Aardsma, Aging: Cause and Cure, 3rd ed. (Loda, IL: Aardsma Research and Publishing, 2023). www.BiblicalChronologist.org.
^ For a map of the discovered route of the Exodus, see, for example, Figure 2 of Gerald E. Aardsma, "The Route of the Exodus, Part XIV: The Location of the Iye-abarim Encampment," The Biblical Chronologist 14.12 (July 16, 2024): 1–2. www.BiblicalChronologist.org. For a new, Bible/science theory of climate change, see Gerald E. Aardsma, "Bible/Science Corrects Mistaken Climate Change Science," The Biblical Chronologist 14.5 (March 21, 2024): 1–22. www.BiblicalChronologist.org and Gerald E. Aardsma, "Bible/Science Corrects Mistaken Climate Change Science Forecasts," The Biblical Chronologist 14.16 (October 31, 2024): 1–8. www.BiblicalChronologist.org.
^ Gerald E. Aardsma, Aging: Cause and Cure, 3rd ed. (Loda, IL: Aardsma Research and Publishing, 2023), 219–221. www.BiblicalChronologist.org.
^ Arts IC, Coolen EJ, Bours MJ, Huyghebaert N, Stuart MA, Bast A, Dagnelie PC. Adenosine 5'-triphosphate (ATP) supplements are not orally bioavailable: a randomized, placebo-controlled cross-over trial in healthy humans. J Int Soc Sports Nutr. 2012 Apr 17;9(1):16. doi: 10.1186 /1550-2783-9-16. PMID: 22510240; PMCID: PMC3441280. pmc.ncbi.nlm.nih.gov/articles/PMC3441280.
^ Schultz MB, Sinclair DA. Why NAD(+) Declines during Aging: It's Destroyed. Cell Metab. 2016 Jun 14;23(6):965-966. doi: 10.1016/j.cmet.2016.05.022. PMID: 27304496; PMCID: PMC5088772. pmc.ncbi.nlm.nih.gov /articles/PMC5088772/.
^ en.wikipedia.org/wiki/Mitochondrial_DNA#mtDNA_in _nuclear_DNA (accessed December 2, 2024).
^ Gerald E. Aardsma, Aging: Cause and Cure, 3rd ed. (Loda, IL: Aardsma Research and Publishing, 2023), chapter 18. www.BiblicalChronologist.org.
^ Vladimir P. Skulachev, Alexander V. Bogachev, and Felix O. Kasparinsky, Principles of Bioenergetics (New York: Springer, 2013), 306–309.
^ Trammell SA, Schmidt MS, Weidemann BJ, Redpath P, Jaksch F, Dellinger RW, Li Z, Abel ED, Migaud ME, Brenner C. Nicotinamide riboside is uniquely and orally bioavailable in mice and humans. Nat Commun. 2016 Oct 10;7:12948. doi: 10.1038/ncomms12948. PMID: 27721479; PMCID: PMC5062546. pmc.ncbi.nlm.nih.gov/ articles/PMC5062546/#b9
^ www.amazon.com/gp/product/B0DHRW3NVY/. This can also be obtained direct from the manufacturer at partiqlar.com/product/pure-nicotinamide-riboside-nr/.